Expression eines Dicarbonsäuretransporters und Löschung von Nebenproduktwegen
Vorherige Einführung des rTCA-Signalwegs in I. orientalis (Stamm SA) (Abb. 1B) ermöglichte die Produktion von SA mit einem Titer von 11.6 g/L in Schüttelkolbenfermentationen13. Um den Titer weiter zu verbessern, haben wir versucht, einen Transporter für SA zu exprimieren. MAE1 für Schizosaccharomyces pombe (SpMAE1) erwies sich als der effizienteste Dicarbonsäuretransporter für den SA-Export18. Das Codon-optimierte SpMAE1 wurde in das Genom des Stammes SA integriert, was zum Stamm SA/MAE1 führte. Die Stämme SA und SA/MAE1 wurden auf SA-Produktion mittels Schüttelkolbenfermentationen in Minimalmedien (SC-URA) mit 50 g/L Glucose unter sauerstofflimitierten Bedingungen untersucht. Die Einführung von SpMAE1 verbesserte den SA-Titer erheblich von 6.8 g/L auf 24.1 g/L (Abb. 2, Ergänzende Abb. 1Aund ergänzende Abb. 1B).
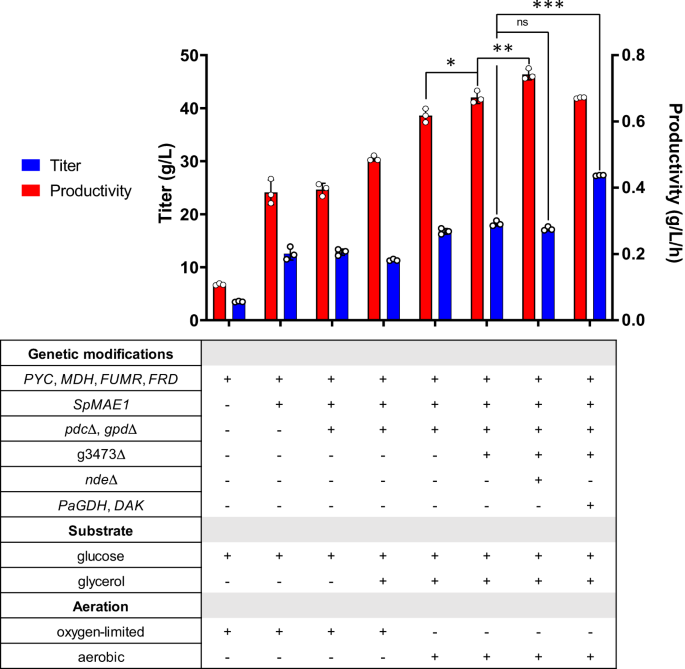
Fehlerbalken, Mittelwert ± SD (n = 3 biologisch unabhängige Proben). *p < 0.05, **p < 0.01, ***p < 0.001, ns nicht signifikant, berechnet durch Two-Tailed Unpaired t-Prüfung. Quelldaten werden als Quelldatendatei bereitgestellt.
Ethanol war das Hauptnebenprodukt und sammelte sich bei der Fermentation des Stammes SA/MAE9.5 mit 1 g/L an. Ethanol entsteht durch die durch Alkoholdehydrogenase (ADH) katalysierte Reaktion, die NADH verwendet, um Acetaldehyd zu Ethanol zu reduzieren. Da die Produktion von SA über den rTCA-Weg NADH erfordert, könnte die Eliminierung des Ethanolbildungswegs die SA-Produktion verbessern. Obwohl sich Glycerin in einer Menge von weniger als 1 g/L anreicherte und nicht das Hauptnebenprodukt war, das bei der Fermentation des Stamms SA/MAE1 beobachtet wurde, kann der durch Glycerin-3-Phosphat-Dehydrogenase (GPD) katalysierte Glycerinbildungsweg potenziell mit dem rTCA-Weg konkurrieren Kohlenstoff und NADH. Also beides PDC (Pyruvatdecarboxylase) und GPD wurden im Stamm SA/MAE1 gelöscht, was zum Stamm SA/MAE1/pdc∆/gpd∆ führte. Beim Löschen von PDC Um die Bildung von Ethanol zu verhindern, sollte der SA-Titer aufgrund der erhöhten Verfügbarkeit von Pyruvat und NADH theoretisch verbessert werden. Die Fermentation des Stamms SA/MAE1/pdc∆/gpd∆ führte unerwartet zu einem ähnlichen SA-Titer von 24.6 g/L und einer Akkumulation von 19.8 g/L Pyruvat unter sauerstofflimitierten Bedingungen (Abb. 2 und ergänzende Abb. 1C).
Kürzlich wurde ein Modell im Genommaßstab erstellt I. orientalisEs wurde vorhergesagt, dass alle ADH-Aktivitäten im Mitochondrium lokalisiert sind19. Somit ist die Entfernung der Ethanolproduktion durch die PDC Die Deletion sollte die zytosolische NADH-Verfügbarkeit nicht erhöhen, was zu keinem Anstieg des SA-Titers führt. Was das zytosolische NADH-Gleichgewicht betrifft, entstehen bei der Glykolyse aus 2 Mol Glucose 2 Mol Pyruvate und 1 Mol NADH, während für die Umwandlung von 1 Mol Pyruvat in 1 Mol Bernsteinsäure 2 Mol NADH erforderlich sind. Obwohl der reduktive TCA-Zyklus theoretisch die höchste Ausbeute aufweist, ist die tatsächliche Ausbeute an SA in Hefen auf nur 1 Mol/Mol Glucose begrenzt. Wir haben dirigiert 13C-Stoffwechselflussanalyse (MFA) und bestätigte, dass der rTCA-Weg den Großteil des durch Glykolyse produzierten zytosolischen NADH effizient für die SA-Produktion nutzte und die Pyruvatausscheidung die Hälfte des im letzten Schritt der Glykolyse produzierten Pyruvats ausmachte (ergänzende Abbildung). 2). Wir exprimierten auch eine zusätzliche Kopie des rTCA-Signalwegs oder eines einzelnen Gens des Signalwegs im Stamm SA/MAE1/pdc∆/gpd∆, um die Kohlenstoffflüsse zwischen Pyruvat und SA weiter zu modulieren, es gab jedoch keine signifikante Änderung des SA-Titers oder der Pyruvatakkumulation (Ergänzende Abb. 3). Daher ist der Mangel an NADH-Versorgung im Zytosol der Hauptengpass für die SA-Produktion über den rTCA-Weg.
Co-Fermentation von Glucose und Glycerin zur SA-Produktion
Da Glukose allein nicht ausreichend zytosolisches NADH für die SA-Produktion produziert, können andere Kohlenstoffquellen in Betracht gezogen werden, um höhere Titer und Ausbeuten zu erzielen. Glycerin hat einen höheren Reduktionsgrad und kann daher mehr reduzierende Äquivalente NADH produzieren als Glucose20,21. Da der Stamm SA/MAE1/pdc∆/gpd∆ Wachstumsdefekte in SC-URA-Medium mit Glycerin als einziger Kohlenstoffquelle aufwies, versuchten wir, die Fermentation dieses Stammes unter Verwendung von SC-URA-Medium mit 50 g/L Glucose und 20 g Glucose durchzuführen /L Glycerin. Zuvor wurde gezeigt, dass die Verwendung von Glucose und Glycerin als duale Kohlenstoffquellen die Umwandlung von Oxalacetat in Malat durch die erhöhte Zufuhr von NADH aus Glycerin in einer technischen Anlage steigert M. succiniciproducens6. Wie in der ergänzenden Abbildung gezeigt. 4A, könnten die Zellen unter sauerstofflimitierten Bedingungen beide Substrate für die SA-Produktion verbrauchen; Allerdings war der Glycerinverbrauch langsam und der SA-Titer verbesserte sich nach 30.5 Tagen Fermentation auf nur 7 g/L. Anschließend führten wir die Fermentation unter aeroben Bedingungen durch und gingen davon aus, dass der Glycerinstoffwechsel unter sauerstofflimitierten Bedingungen eingeschränkt sein könnte. Unter aeroben Bedingungen wurden sowohl Glukose als auch Glycerin schneller verbraucht, was die Produktion von 38.6 g/L SA ermöglichte (Abb. 2 und ergänzende Abb. 4B). Wir haben auch die Fermentation des Stammes SA/MAE1/pdc∆/gpd∆ unter Verwendung von SC-URA-Medium mit 50 g/L Glucose unter aeroben Bedingungen getestet. Während der rTCA-Weg ein fermentativer Weg ist und eine höhere Belüftung möglicherweise mehr Kohlenstofffluss in den TCA-Zyklus für die aerobe Atmung leiten könnte, beobachteten wir interessanterweise, dass aerobe Bedingungen im Vergleich zu sauerstofflimitierten Bedingungen zu ähnlichen Titern führten und die Zellen in der Lage waren, Glukose zu verbrauchen schneller, was zu höheren Produktivitäten führt (ergänzende Abb. 4C).
Deletionen eines Dicarbonsäureimporteurs und einer externen NADH-Dehydrogenase
Anschließend wurden weitere Gendeletionen versucht, um die SA-Produktion zu steigern. Kürzlich wurden die Carboxylattransporter PkJEN2-1 und PkJEN2-2 der JEN-Familie eingeführt Pichia kudriavzevii Es wurde charakterisiert, dass sie an der Aufnahme von Dicarbonsäuren beteiligt sind22,23. PkJEN2-1 und PkJEN2-2 wurden als g3473 und g3068 in kommentiert I. orientalis, jeweils. g3473 wurde aus dem Stamm SA/MAE1/pdc∆/gpd∆ gelöscht, was zum Stamm g3473∆ führte. Die Fermentation dieses Stammes in SC-URA-Medium mit 50 g/L Glucose und 20 g/L Glycerin verbesserte den SA-Titer auf 42.0 g/L (Abb. 2 und ergänzende Abb. 5A), was darauf hindeutet, dass die Verhinderung des erneuten Eindringens von SA in die Zellen von Vorteil war. g3068 wurde im Stamm g3473∆ weiter ausgeschaltet; Wir beobachteten jedoch, dass die Störung beider JEN2-Transporter den SA-Titer auf 34.5 g/l senkte und somit keinen Nutzen brachte (ergänzende Abbildung). 5B). Dieses Ergebnis stimmte nicht mit dem vorherigen Bericht überein, wonach beide JEN-Transporter gestrichen wurden P. kudriavzevii CY902 führte zu einem höheren SA-Titer als einzelne Gendeletionen, was möglicherweise auf unterschiedliche genetische Hintergründe zurückzuführen ist. P. kudriavzevii CY902 wurde so konstruiert, dass es SA über den oxidativen TCA (oTCA)-Weg durch Deletion des Gens der Succinat-Dehydrogenase-Komplex-Untereinheit produziert SDH5, während SA-Produktion in unserem entwickelt I. orientalis SD108 wurde über den rTCA-Weg erreicht. Basierend auf MFA wurde außerdem eine kleine Menge zytosolisches NADH durch die externe mitochondriale NADH-Dehydrogenase (NDE) oxidiert, die Elektronen vom zytosolischen NADH zur mitochondrialen Elektronentransportkette transportiert (ergänzende Abbildung). 2). NTE wurde auf eine Störung im Stamm g3473∆ abzielt, was zum Stamm g3473∆/nde∆ führte. Im Vergleich zum Stamm g3473∆, NTE Die Löschung verbesserte den SA-Titer weiter auf 46.4 g/L, was auf den Knockout von hindeutet NTE erhöhte den zytosolischen NADH-Pool für die Produktion von SA (Abb. 2 und ergänzende Abb. 5C). Dennoch ist die Störung von NTE senkte die Glukoseverbrauchsrate; Daher hatte der Stamm g3473∆/nde∆ trotz seines höheren Titers eine ähnliche Produktivität wie der Stamm g3473∆ (Abb. 2).
Verbesserung des Glycerinverbrauchs
Der langsame Glycerinverbrauch deutete darauf hin, dass der endogene Glycerinstoffwechsel möglicherweise nicht sehr aktiv ist. Zuvor war eine Überexpression der Glycerindehydrogenase (GDH) aus Pichia angusta und endogene Dihydroxyacetonkinase (DAK) etablierte einen NADH-produzierenden Glycerin-Verbrauchsweg in S. cerevisiae24. Daher versuchten wir, eine ähnliche Strategie anzuwenden, um den Glycerinverbrauch in zu verbessern I. orientalis. Das Codon optimiert PaGDH und endogen DAK wurden in den Stämmen g3473∆ und g3473∆/nde∆ überexprimiert, was zu den Stämmen g3473∆/PaGDH-DAK bzw. g3473∆/nde∆/PaGDH-DAK führte. Fermentationen dieser Stämme in SC-URA-Medium mit 50 g/L Glucose und 20 g/L Glycerin führten nicht zu höheren SA-Titern; g3473∆/PaGDH-DAK und g3473∆/nde∆/PaGDH-DAK produzierten SA mit Titern von 41.9 g/L bzw. 46.5 g/L, ähnlich den Titern, die von den Elternstämmen ohne Überexpression von erreicht wurden PaGDH und DAK (Feige. 2, Ergänzende Abb. 6Aund ergänzende Abb. 6B). Allerdings ist die Überexpression von PaGDH und DAK wirkte sich sowohl auf die Glukose- als auch auf die Glycerinverwertungsrate positiv aus. Die Produktivitäten stiegen von 0.29 auf 0.44 g/L/h im Stamm g3473∆/PaGDH-DAK und von 0.28 auf 0.32 g/L/h im Stamm g3473∆/nde∆/PaGDH-DAK (Abb. 2 und ergänzende Abb. 6C).
Der Stamm g3473∆/PaGDH-DAK könnte bei der Fermentation mit 25.4 g/L Glucose 50 g/L SA produzieren, während aus 41.9 g/L Glucose und 50 g/L Glycerin 20 g/L SA gewonnen werden könnten (Ergänzung). Feige. 7A). Da der SA-Titer von 41.9 g/L auch allein durch die einfache Verwendung von mehr anfänglicher Glukose in der Fermentation unter alleiniger Verwendung von Glukose erreicht werden könnte, kann man die Vorteile der Verwendung von Glukose und Glycerin als duale Kohlenstoffquellen in Frage stellen. Auf Kohlenstoffäquivalentbasis entspricht 1 Gramm Glucose 1 Gramm Glycerin. Die Verwendung von 50 g/L Glucose und 20 g/L Glycerin ermöglichte eine SA-Ausbeute von 0.60 g/g Glucoseäquivalent, was höher war als die Ausbeute von 0.51 g/g Glucose aus der Fermentation mit nur 50 g/L Glucose (ergänzende Abbildung). . 7B). Darüber hinaus konnte der Stamm g70∆/PaGDH-DAK aus 50 g/L Glucose, einer Konzentration, die 20 g/L Glucose und 3473 g/L Glycerin entspricht, SA mit einem Titer von nur 35.6 g/L und einer Ausbeute von 0.50 produzieren g/g Glukose (Ergänzende Abb. 7). Daher ermöglichte die Verwendung einer Mischung aus Glucose und Glycerin als Kohlenstoffquellen die SA-Produktion mit höheren Titern und höheren Ausbeuten als bei Verwendung einer entsprechenden Menge Glucose. Wir verglichen auch die SA-Produktion im Stamm g3473∆/PaGDH-DAK unter Verwendung von 50 g/L Glucose und verschiedenen Glycerinkonzentrationen von 10 g/L, 20 g/L und 30 g/L (ergänzende Abbildung). 8). Wie erwartet stieg der SA-Titer mit zunehmender anfänglicher Glycerinmenge; Daher war ein Vergleich zwischen den Titern nicht aussagekräftig. Außerdem war die Ausbeute am niedrigsten, wenn 10 g/L Glycerin verwendet wurden. Bei der Verwendung von 20 g/L und 30 g/L Glycerin wurde kein signifikanter Ertragsunterschied beobachtet; Die Produktivität war jedoch am höchsten, wenn 20 g/L Glycerin verwendet wurden. Daher hielten wir 20 g/L Glycerin für die optimale Konzentration zur Verwendung mit 50 g/L Glukose für die SA-Produktion.
Wir haben auch versucht, die katabolische Unterdrückung von Glukose beim Glycerinverbrauch durch die Deletion einer Hexokinase zu mildern, die nachweislich die Glukosephosphorylierungsrate verringert und die gemeinsame Nutzung von Glukose und Xylose ermöglicht S. cerevisiae25. Durch BLAST-Analyse wurden drei potenzielle Hexokinase-Gene (g1398, g2945 und g3837) bestimmt, und nur die Deletion von g3837 im Stamm g3473∆/PaGDH-DAK ermöglichte den gleichzeitigen Verbrauch von Glucose und Glycerin (ergänzende Abb. 9). Während ähnliche SA-Titer erreicht werden konnten, verringerte die Deletion von g3837 die Glukose- und Glycerinverbrauchsraten, was zu keiner Steigerung der Produktivität führte.
Fed-Batch-Fermentationen und Scale-up
Im Anschluss an die Fermentationen im Schüttelkolben führten wir Fed-Batch-Fermentationen durch, um den SA-Titer zu erhöhen und die Leistung unseres gentechnisch veränderten Stamms in der Großproduktion zu bewerten. Um die überlegene Toleranz gegenüber niedrigem pH-Wert auszunutzen I. orientalisWir entschieden uns dafür, die Fed-Batch-Fermentationen bei pH 3 durchzuführen. Bei diesem pH-Wert sind etwa 90 % der SA-Spezies vollständig protoniertes SA, während die restlichen 10 % der Spezies Hydrogensuccinat sind26. Wir haben zunächst die Leistung des Stammes g3473∆/PaGDH-DAK getestet, der aufgrund der höheren Produktivität dem Stamm g3473∆/nde∆/PaGDH-DAK vorgezogen wurde, indem wir SC-URA-Medium mit 50 g/L Glucose und 20 g/L Glucose verwendet haben Glycerin in der Batch-Fermentation in einem Tischbioreaktor mit einer Größe von 0.3 l und einem Arbeitsvolumen von 0.1 l unter statischen Bedingungen des Rührens und kontinuierlicher Einleitung von O2 und CO2. Wir beobachteten, dass die Titer (27.1 g/L und 30.7 g/L bei 0.333 vvm (Volumen pro Arbeitsvolumen pro Minute) CO2 und 0.667 vvm CO2) waren viel niedriger als der bei der Schüttelkolbenfermentation erhaltene Titer (42.1 g/L) (ergänzende Abb. 10A,B). Während sowohl im Reaktor als auch im Schüttelkolben ähnliche SA-Titer aus Glucose hergestellt werden konnten, waren die während der Glycerinnutzungsphase erzeugten SA-Titer im Bioreaktor viel niedriger. Wir führten auch eine Batch-Fermentation im Bioreaktor mit dem Stamm g3473∆/PaGDH-DAK/g3837∆ durch und beobachteten, dass dieser Stamm während der Glycerinverbrauchsphase mehr SA produzieren konnte und ein Titer von 38.8 g/L SA bei 0.167 vvm O erhalten werden konnte2 und 0.667 vvm CO2 (Ergänzende Abb. 10C, D). Wir gingen davon aus, dass bei der Verwendung von Glycerin in der Bioreaktorumgebung mit höherer Belüftung als in der Schüttelkolbenumgebung möglicherweise mehr Kohlenstofffluss in den TCA-Kreislauf geleitet wird und zu einem niedrigeren SA-Titer führt. Andererseits könnte die Deletion von g3837 die Aktivität der TCA-Zyklus-Gene unterdrücken und die SA-Produktion verbessern. Mithilfe einer Echtzeit-PCR-Analyse wurden die Transkriptionsniveaus von Genen im rTCA-Signalweg und einigen ausgewählten Genen im TCA-Zyklus (Citratsynthase, CIT; Aconitase, ACO; und Isocitratdehydrogenase, HDI) in den Stämmen g3473∆/PaGDH-DAK mit oder ohne g3837-Deletion, gezüchtet in YP-Medium mit Glycerin. Wir beobachteten, dass das Knockout von g3837 ähnliche Expressionen von Genen im rTCA-Signalweg aufrechterhielt, jedoch die Expressionsniveaus von senkte CIT, ein Homolog von ACO und HDIs (Ergänzende Abb. 11). Daher könnten die geringeren Aktivitäten von Genen im TCA-Zyklus zu einem höheren SA-Titer führen, der vom Stamm g3473∆/PaGDH-DAK/g3837∆ im Bioreaktor erhalten wird. Die Fed-Batch-Fermentation des Stammes g3473∆/PaGDH-DAK/g3837∆ in SC-URA-Medium mit Glukose- und Glycerinfütterung ergab 109.5 g/l SA mit einer Ausbeute von 0.65 g/g Glukoseäquivalent und einer Produktivität von 0.54 g/l. L/h (Abb. 3A). Am Ende der Fermentation beobachteten wir die Bildung von Kristallen, was wahrscheinlich SA war (ergänzende Abbildung). 12). Während andere organische Säuren wie Milch- und Essigsäure in wässriger Brühe bei pH 1–14 vollständig mischbar sind, nimmt die Löslichkeit von SA ab, wenn der pH-Wert saurer wird27.
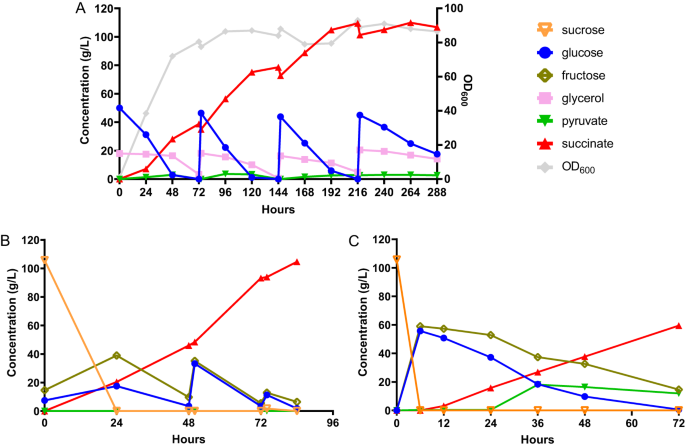
A Fed-Batch-Fermentation des Stammes g3473∆/PaGDH-DAK/g3873∆ in Minimalmedium mit Glucose und Glycerin. B Fed-Batch-Fermentation des Stammes g3473∆/PaGDH-DAK/ScSUC2 in Zuckerrohrsaftmedium. C Batch-Fermentation des Stammes g3473∆/PaGDH-DAK/ScSUC2 in Zuckerrohrsaftmedium in einem Reaktor im Pilotmaßstab. Quelldaten werden als Quelldatendatei bereitgestellt.
Im Anschluss an die hohe fermentative Leistung unseres Rekombinanten I. orientalis Anschließend testeten wir die Produktion von SA mit einem echten Industriesubstrat, Zuckerrohrsaft. Zuckerrohr ist die energieeffizienteste mehrjährige C4-Pflanze und weist im Vergleich zu anderen Nutzpflanzen wie Rutenhirse und Chinaschilf einen höheren Biomasseertrag auf28. Darüber hinaus ist Zuckerrohrsaft als saccharosebasierter Rohstoff günstiger als Glukose- und stärkebasierte Substrate wie Mais und Maniok29. Da I. orientalis ist nicht in der Lage, Saccharose, die Invertase, zu verwerten SUC2 für S. cerevisiae wurde in g3473∆/PaGDH-DAK ausgedrückt. Die Batch-Fermentation fand in den ersten 48 Stunden statt und SA konnte mit einem Titer von 46.0 g/L, einer Ausbeute von 0.40 g/g Glucoseäquivalent und einer Produktivität von 0.96 g/L/h produziert werden. Durch die anschließende Zufuhr von konzentriertem Zuckerrohrsaft konnte unser gentechnisch veränderter Stamm SA mit einem Titer von 104.6 g/L, einer Ausbeute von 0.63 g/g Glucoseäquivalent und einer Produktivität von 1.25 g/L/h im Labormaßstab produzieren (Abb. 3B).
Darüber hinaus haben wir unseren SA-Fermentationsprozess mit Zuckerrohrsaft vom Labormaßstab auf einen Pilotmaßstab erweitert. Hier haben wir die Prozessparameter bestimmt, um eine ähnliche Leistungsaufnahme pro Volumeneinheit und die gleiche Reynolds-Zahl zwischen Bioreaktoren im Labormaßstab und im Pilotmaßstab aufrechtzuerhalten, und die Batch-Fermentation wurde in einem Bioreaktor im Pilotmaßstab mit einer Größe von 75 l und einem Arbeitsvolumen durchgeführt Volumen von 30 l bzw. ein 300-facher Skalierungsfaktor im Vergleich zum Tisch-Bioreaktor. Unser Stamm konnte SA mit einem Titer von 63.1 g/L, einer Ausbeute von 0.50 g/g Glucoseäquivalent und einer Produktivität von 0.66 g/L/h bei pH 3 produzieren (Abb. 3C). Aufgrund des Volumenbedarfs haben wir keine Fed-Batch-Fermentation im Pilotmaßstab versucht; Dennoch waren unser Titer und unsere Ausbeute für die Batch-Fermentation im Bioreaktor im Pilotmaßstab vergleichbar mit denen in den Bioreaktoren im Labormaßstab. Daher erwarteten wir eine ähnliche Leistung des Stammes bei der Fed-Batch-Fermentation im Pilotmaßstab.
Wir haben den gesamten Produktionsprozess von SA weiter vervollständigt, indem wir einen DSP zur Gewinnung von SA aus Zuckerrohrsaft-Fermentationsbrühe mithilfe einer zweistufigen Vakuumdestillation und -kristallisation entwickelt haben. Ohne weitere Ansäuerung der aus der Fermentation im Pilotmaßstab erhaltenen Fermentationsbrühe mit 63.1 g/L SA betrug die maximale Ausbeute in der ersten Stufe 31.0 %. Anschließend wurde das Filtrat aus Stufe 1 mittels Vakuumdestillation auf 50 % seines Volumens eingeengt und der zweiten Stufe der Kristallisation unterzogen. Die Ausbeute an SA aus Stufe 2 betrug 47.7 %, und für beide Stufen wurden ähnliche Mengen an SA-Kristallen erhalten (1.98 g für Stufe 1 und 2.10 g für Stufe 2). Somit wurde durch zweistufige Vakuumdestillation und Kristallisation eine SA-Gesamtausbeute von 64.0 % aus der Fermentationsbrühe mit niedrigem pH-Wert erzielt. Darüber hinaus wurde die Reinheit der in Stufe 1 und Stufe 2 gewonnenen SA-Kristalle auf 88.9 % bzw. 86.23 % geschätzt. Das erzielte Ergebnis stimmte mit den Ergebnissen einer früheren Studie überein30. Aus den Ergebnissen ging hervor, dass die Kristallisation von SA in hoher Reinheit (>85 %) aus der unbehandelten Fermentationsbrühe erfolgreich war. Es sind jedoch weitere Untersuchungen erforderlich, um die färbenden Verunreinigungen zu beseitigen und SA-Kristalle in kommerzieller Qualität zu gewinnen.
Technoökonomische Analyse und Ökobilanz
Wir haben End-to-End-Bioraffinerien entworfen und simuliert, die in der Lage sind, Zuckerrohr als Ausgangsmaterial zu akzeptieren, es zu Zuckerrohrsaft (Saccharose, Glucose und Fructose) zu verzuckern und den Zucker mithilfe von SA zu SA zu fermentieren I. orientalisund Abtrennen der Fermentationsbrühe, um getrocknete SA-Kristalle zu gewinnen (ergänzende Abb. 13) mit einer jährlichen Produktionskapazität von 26,800 Tonnen SA (die weltweite Nachfrage nach SA lag 2013 bei etwa 76,000 Tonnen).31). Die Bioraffinerien wurden unter alternativen Fermentationsszenarien simuliert, wobei Annahmen für Ausbeute, Titer und Produktivität der Fermentationsleistung entsprachen, die in Experimenten im Batch-Modus (Labor-Batch-Szenario) und im Fed-Batch-Modus (Labor-Fed-Batch-Szenario) erzielt wurde die Einrichtung des Batch-Modus im Pilotmaßstab (Pilot-Batch-Szenario). Um die finanzielle Rentabilität und die Umweltvorteile der entwickelten SA-Pfade zu charakterisieren, führten wir für jedes Szenario eine TEA und eine Ökobilanz unter Basisannahmen sowie unter Unsicherheit durch (2000 Monte-Carlo-Simulationen für jedes Szenario mit lateinischer Hypercube-Stichprobe; die Verteilung der Ergebnisse bei alternativen Zahlen). der Monte-Carlo-Simulationen ist in der Ergänzungstabelle aufgeführt 1 und die angenommenen Basiswerte und Verteilungen aller unsicheren Parameter für jedes Szenario werden in den Zusatzdaten angegeben 1). Wir haben den Mindestverkaufspreis des Produkts (MPSP, im Jahr 2016 $ mit einer internen Rendite von 10 %) und das 100-jährige globale Erwärmungspotenzial (GWP) verwendet100; (Cradle-to-Grave) und fossiler Energieverbrauch (FEC; Cradle-to-Gate) als Messgrößen zur Darstellung der TEA- und LCA-Ergebnisse. Wir haben auch Sensitivitätsanalysen mithilfe der Rangkorrelationskoeffizienten nach Spearman (Spearmans ρ) durchgeführt, um die Haupttreiber der Produktionskosten und Umweltauswirkungen zu identifizieren. Um schließlich Ziele für weitere Verbesserungen der finanziellen Rentabilität und der ökologischen Nachhaltigkeit festzulegen und zu priorisieren, haben wir Bioraffinerien für die gesamte potenzielle Fermentationsleistungslandschaft entworfen und simuliert (d. h. jeweils 2500 Ertrag-Titer-Kombinationen für eine Reihe von Produktivitäten sowohl für die neutrale als auch für die Fermentation mit niedrigem pH-Wert). ).
Basierend auf der experimentellen Leistung im Labor-Batch-Szenario könnte die Bioraffinerie SA mit einem geschätzten MPSP von 1.70 $/kg produzieren (Basislinie; Abb. 4A) mit einer Spanne von 1.51–1.92 $/kg (5.–95. Perzentil; im Folgenden in Klammern). Das GWP der Bioraffinerie100 und FEC wurden in diesem Szenario auf 1.95 kg CO geschätzt2-eq./kg (1.37–2.65 kg CO2-eq./kg) bzw. -3.74 MJ/kg (−12.9–5.39 MJ/kg) (Abb. 4B und ergänzende Abb. 14A). Im Labor-Fed-Batch-Szenario (mit verbessertem SA-Titer, Ertrag und Produktivität der Fermentation im Vergleich zum Labor-Batch-Szenario) betrug der MPSP der Bioraffinerie 1.06 $/kg (0.96–1.22 $/kg), GWP100 betrug 0.93 kg CO2-eq./kg (0.71–1.32 kg CO2-eq./kg) und FEC betrug −5.36 MJ/kg (−8.97–0.213 MJ/kg). Im Pilot-Batch-Szenario (mit verbesserter SA-Ausbeute und Titer der Fermentation im Vergleich zum Labor-Batch-Szenario, aber niedriger als beim Fed-Batch-Labor-Szenario und geringerer Produktivität als bei beiden Labor-Szenarios) hatte die Bioraffinerie einen geschätzten MPSP von 1.37 $/kg (1.23–1.54 $/kg), GWP100 von 1.67 kg CO2-eq./kg (1.22–2.17 kg CO2-eq./kg) und FEC von −0.21 MJ/kg (−7.08 bis 6.47 MJ/kg). Ein Sankey-Diagramm, das den Kohlenstofffluss durch die Bioraffinerie für dieses Szenario darstellt, wurde auch in der ergänzenden Abbildung gezeigt. 15.
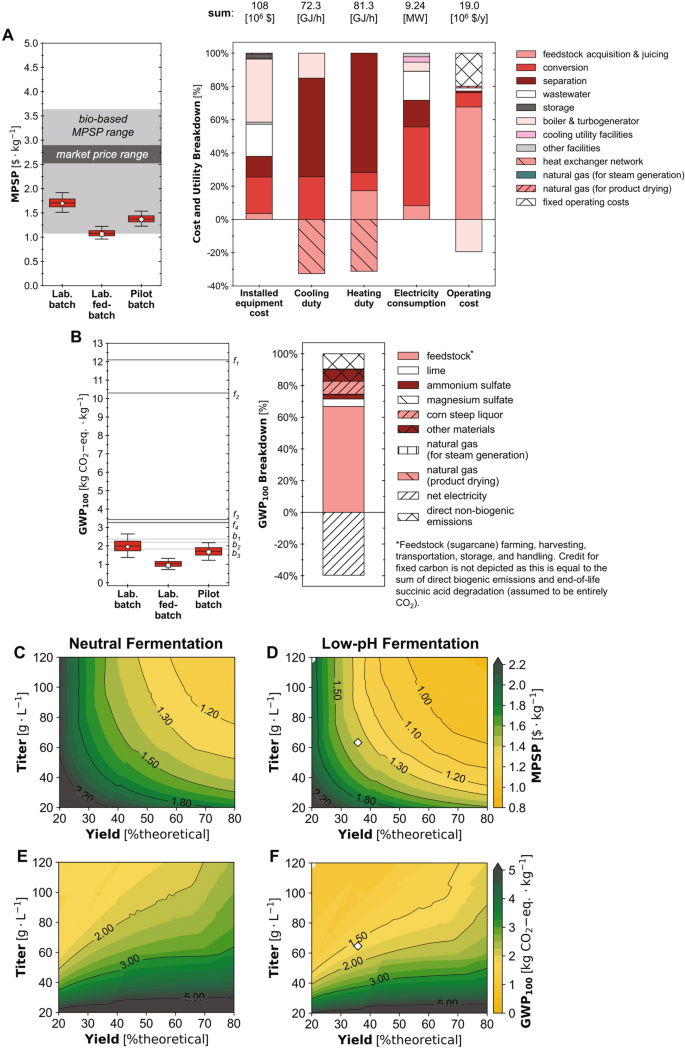
Unsicherheiten (Box- und Whisker-Diagramme) und Aufschlüsselungen (gestapelte Balkendiagramme) für (A) Mindestproduktverkaufspreis (MPSP) und (B) 100-jähriges globales Erwärmungspotenzial (GWP) von der Wiege bis zur Bahre100). Whiskers, Kästchen und die Mittellinie stellen das 5./95., 25./75. und 50. Perzentil aus Monte-Carlo-Simulationen aus dem Jahr 2000 dar (n = 2000 Simulationen) für jedes Szenario. Fünfeckige, quadratische und rautenförmige Markierungen stellen Basisergebnisse für die Szenarios Laborcharge (Lab. Batch), Labor-Fed-Batch (Lab. Fed-Batch) und Pilotcharge (Pilot-Batch) dar. Gestapelte Balkendiagramme zeigen Basisergebnisse für das Pilot-Batch-Szenario. Ergebnisse für andere Szenarien sind im SI enthalten. Der Stromverbrauch umfasst nur den Verbrauch der Anlage; Aus Gründen der Übersichtlichkeit wurde die Produktion in der dargestellten Aufschlüsselung ausgeschlossen. Tabellarische Aufschlüsselungsdaten für Material- und Installationskosten, Heiz- und Kühlleistungen, Stromverbrauch, GWP100, und FEC sind online verfügbar62. Markierte dunkelgraue Linien kennzeichnen gemeldete Auswirkungen für fossilbasierte Produktionswege (f152; f2-f451). Markierte hellgraue Linien kennzeichnen gemeldete Auswirkungen für alternative biobasierte Produktionswege (b1).51; b253; b352). Wo GWP100 wurde als „Cradle-to-Gate“ gemeldet, 1.49 kg CO2-Äquivalent/kg wurden als Auswirkungen am Lebensende hinzugefügt, um der Übereinstimmung mit dieser Studie und Dunn et al. 2015. Die Werte für alle gemeldeten MPSPs und Auswirkungen vor und nach der Anpassung sind in der Ergänzungstabelle aufgeführt 3 und 4. (C, D) MPSP und (E, F) GWP100 über 2500 Fermentationsausbeute-Titer-Kombinationen bei der Basisproduktivität des Pilotchargen-Szenarios (0.66 g/l/h) für Neutral (linkes Feld; C, E) und niedriger pH-Wert (rechtes Feld; D, F) Gärung. Die Ausbeute wird als % des theoretischen Maximums (% theoretisch) angezeigt, skaliert auf die theoretische Maximalausbeute von 1.31 g/g-Glukoseäquivalent (basierend auf der Kohlenstoffbilanz). Für einen bestimmten Punkt in der Abbildung gilt: xDer Wert auf der Achse stellt die Ausbeute dar, der Wert auf der Y-Achse stellt den Titer dar und die Farb- und Konturlinien stellen den Wert von MPSP und GWP dar100. Diamantmarkierungen zeigen Basisergebnisse für das Pilotchargen-Szenario. Quelldaten werden als Quelldatendatei bereitgestellt.
Bei den 28 Parametern, denen die Unsicherheit für das Pilotchargen-Szenario zugeschrieben wurde, stellten wir fest, dass MPSP am empfindlichsten auf die Fermentations-SA-Ausbeute reagierte (Spearmans ρ von –0.60; alle Unsicherheitsverteilungen sind in den ergänzenden Daten aufgeführt 1 und Spearmans ρ-Werte für alle Parameter sind in der Ergänzungstabelle angegeben 2). MPSP reagierte auch erheblich empfindlich auf den Stückpreis für Futterzuckerrohr (Spearmans ρ von 0.39), die Betriebszeit der Anlage (–0.38), die Kapazität der Anlage für Futterzuckerrohr (–0.31) und den Fermentations-SA-Titer (–0.30). GWP100 reagierte am empfindlichsten auf den Kesselwirkungsgrad (Spearmans ρ von −0.63). GWP100 war auch signifikant empfindlich gegenüber dem SA-Titer und der Ausbeute der Fermentation, mit Spearmans ρ-Werten von –0.62 bzw. 0.32. FEC reagierte am empfindlichsten auf die Fermentations-SA-Ausbeute (Spearman-ρ von 0.63) und auch empfindlich auf den Kesselwirkungsgrad (–0.56) und den Fermentations-SA-Titer (–0.49). Um die Auswirkungen der Fermentationsleistung weiter zu charakterisieren, führten wir TEA und LCA in der gesamten Fermentationsleistungslandschaft durch (Abb. 4C – F und ergänzende Abb. 14B, C) und simuliert 2500 Ertrags-Titer-Kombinationen über eine Reihe von Produktivitäten für jedes von zwei alternativen Regimen: Fermentation mit niedrigem pH-Wert (d. h. mit Fermentation bei einem pH-Wert von 3, der durch Basenzugabe während der Fermentation kontrolliert wird und nach der Fermentation keine Ansäuerung erforderlich ist) und neutrale Fermentation (d. h. mit vollständiger Neutralisierung von SA durch Basenzugabe während der Fermentation und vollständiger Wiederansäuerung nach der Fermentation). Die Ergebnisse für ein alternatives Szenario mit niedrigem pH-Wert und erforderlicher erneuter Ansäuerung nach der Fermentation sind in der ergänzenden Abbildung dargestellt. 16.
- SEO-gestützte Content- und PR-Distribution. Holen Sie sich noch heute Verstärkung.
- PlatoData.Network Vertikale generative KI. Motiviere dich selbst. Hier zugreifen.
- PlatoAiStream. Web3-Intelligenz. Wissen verstärkt. Hier zugreifen.
- PlatoESG. Kohlenstoff, CleanTech, Energie, Umwelt, Solar, Abfallwirtschaft. Hier zugreifen.
- PlatoHealth. Informationen zu Biotechnologie und klinischen Studien. Hier zugreifen.
- Quelle: https://www.nature.com/articles/s41467-023-41616-9

