El L álbum es una conocida fuente de lignanos. Se han realizado varios estudios sobre sus propiedades fisiológicas, bioquímicas y expresión de algunos genes clave implicados en la biosíntesis de lignanos técnicas de cultivo in vitro3,9,21. Este estudio tuvo como objetivo reconocer una base común de cambios transcripcionales, especialmente cambios asociados con la biosíntesis de PTOX que ocurren en diferentes órganos durante múltiples estreses en cultivos in vivo.
Los perfiles de expresión mostraron obviamente diferencias en el número de DEG regulados hacia arriba y hacia abajo bajo múltiples tensiones. Se observaron menos DEG en el estrés de esta planta relacionado con las raíces (como la toxicidad del Al) en comparación con los brotes. Estas diferencias están impulsadas por diferencias entre conjuntos de datos que pueden tener varias fuentes, como la variabilidad biológica o técnica.
La identificación de una gran cantidad de DEG bajo estrés por sequía indicó que una parte considerable del transcriptoma estaba afectado por este estrés.22. Bajo estrés por sequía, las diferencias entre los genotipos tolerantes y susceptibles surgen en los patrones de expresión génica y los fenotipos.10. Además, se ha observado que el nivel de expresión de los genes de respuesta al estrés suele ser mayor en los genotipos de tolerancia.13. Por lo tanto, estos resultados corroboran el concepto de que el lino tiene mejor tolerancia a la sequía que otros estreses.10.
De acuerdo con los DEG identificados, 20 genes comúnmente regulados que responden a estreses bióticos y abióticos y 38 genes comúnmente regulados que responden a estreses abióticos se superpusieron utilizando análisis de diagrama de Venn en el brote y la raíz, respectivamente. Entonces, solo un gen comúnmente regulado, la endoquitinasa EP3, se identificó con un aumento significativo en ambos órganos. El EP3, que cataliza la hidrólisis de la quitina, participa en los procesos de desarrollo de las plantas, la generación de moléculas señalizadoras, las respuestas de defensa de las plantas y la muerte celular programada23,24. La expresión de la EP3 aumento del gen en respuesta a la sequía, la salinidad, los tratamientos con luz ultravioleta, el tratamiento con inductores exógenos, las heridas y el ataque de patógenos11,23,24. Además, el gen de la β-1,3-glucanasa aumentó en los resultados relacionados con el brote, que previamente se demostró que es crucial para la resistencia del lino a Fusarium spp.25. Las plantas de lino con sobreexpresión de β-1,3-glucanasa, generando inhibidores de pectinasa y empleando quitinasas y peroxidasas aumentaron la producción de SM y cambiaron la composición de la pared celular, lo que condujo a la construcción de una barrera para el crecimiento de hongos26.
Los procesos biológicos, incluidos el metabolismo secundario, el estrés, el desarrollo, la pared celular, el metabolismo de los lípidos y la degradación de las proteínas, se vieron comúnmente alterados por todos los tipos de estrés. Tal regulación común refleja la flexibilidad de los sistemas biológicos a través del ajuste de redes metabólicas complejas en respuesta a estimulantes durante la evolución de la vida inmóvil de la planta.11.
Los términos GO comunes que responden al estrés consistieron en familias de factores de transcripción AP2/ERF, HB-7 y NAC que aumentaron significativamente en el tratamiento con ABA y el estrés por sequía y disminuyeron en Fusarium tratamiento y k+ deficiencia. El estudio del estrés individual ha identificado 11 TF principales, incluidos bHLH (hélice-bucle-hélice básica), C2H2, NAC, MYB, ERF, bZIP, WRKY, MYB, DREB, HSF y NFYA10 como principales reguladores conocidos de las vías de resistencia abiótica. bajo sequía repetida en lino10. Los TF de NAC también contribuyeron a la respuesta de estrés a la tolerancia del maíz, el arroz y el lino al estrés por aluminio. Estos TF, junto con MADS-box, ajustan el crecimiento y desarrollo de las plantas e involucran la alteración de la pared celular que conduce a la tolerancia al aluminio.13. Se ha demostrado que la mayoría de los TF relacionados con la falta de potasio pertenecen a MYB, bHLH, NAC, B3, bZIP, WRKY y AP2/ERF, y participan en los procesos fisiológicos de las plantas, la resistencia al estrés y el metabolismo secundario.18. Las vías de señalización del estrés comparten componentes comunes que comprenden cascadas de ROS, iones de calcio, hormonas, TF y proteína quinasa activada por mitógeno (MAPK).27.
Los términos raíz GO mostraron que los procesos biológicos, a saber, la transformación de TCA/org, la actividad de la oxidorreductasa, la pared celular, el transporte, el metabolismo hormonal y la señalización de la quinasa del receptor, se modulan específicamente bajo estrés abiótico. En un estudio de lino bajo estrés individual, los términos GO de la actividad de la oxidorreductasa, particularmente las peroxidasas, la pared celular, la homeostasis iónica y la respuesta al estrés cambiaron más en condiciones desfavorables de pH y deficiencia de Zn.12.
Algunos de nuestros resultados coincidieron con los estudios individuales, mientras que otros se opusieron. Estos factores estresantes individuales pueden causar diferentes respuestas de ajuste, que comprenden varios componentes compartidos en las plantas. Cuando las plantas se encuentran simultáneamente con combinaciones de estrés, podrían requerir respuestas moleculares, fisiológicas y metabólicas similares u opuestas. La elección precisa de qué estrategia de ajuste durante el estrés múltiple presumiblemente se verá afectada por factores como la intensidad de cada estrés individual, el curso temporal del estrés, la edad de la planta y el genotipo (tolerante o susceptible a cualquiera de los estrés individuales)28. El tipo de ajuste puede influir en la acumulación de SM, provocando la modulación espacial y temporal de las vías biosintéticas, mejorando la posibilidad de supervivencia en entornos estresantes a largo plazo.29.
El análisis de MapMan clasificó los SM en 16 grupos. Las vías de fenilpropanoide y flavonoide se vieron muy afectadas por todos los estreses. La mayor abundancia de transcritos se relacionó con las vías de flavonoides y fenilpropanoides bajo estrés por sequía. El aumento de estos compuestos estuvo muy relacionado con el balance de carbohidratos entre fuentes y sumideros. Además, se ha informado que los potenciales hídricos disminuyeron en la planta bajo estrés severo por sequía, lo que llevó al transporte de azúcares solubles. Por lo tanto, la acumulación de flavonoides y fenoles aumentó30,31. Los resultados relacionados con el estrés por sequía en este trabajo mostraron una reducción de la abundancia de transcritos cinco veces (-5) en la vía de los glucosinolatos. De acuerdo con estos resultados, el estrés por sequía redujo significativamente el contenido de glucosinolatos en Boechera holboellii Hornem. Á.Löve & D.Löve y algunos Brassica carinata cultivares A.Braun, mientras que algunos B. carinata cultivares mostraron un aumento significativo en glucosinolatos32,33. El tratamiento con sulfato de potasio redujo el contenido de glucosinolatos durante condiciones de sequía en la canola en comparación con las plantas no tratadas34. Las raíces mostraron algunos cambios en la abundancia de transcritos de SM. Sin embargo, las transcripciones sensibles al estrés se alteran principalmente en condiciones adversas. Los SM, como el ácido sinápico, la lignina y los flavanoles, con funciones defensivas, aumentaron en Sinopodophyllum hexandrum Royle bajo déficit de agua35.
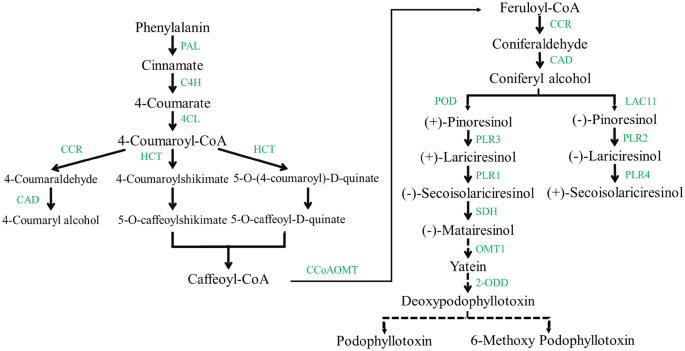
Varios compuestos fenólicos, incluidos flavonoides, monolignoles, lignanos, ligninas, cumarinas y taninos hidrolizables, se forman a través de la ruta de los fenilpropanoides.36. Linum género, particularmente L álbum, contiene los niveles más altos de lignanos, especialmente PTOX. Sin embargo, muchos estudios han investigado la ruta de biosíntesis de lignanos, la ruta de lignanos hasta que el producto final PTOX no se ha aclarado por completo.8,20.
Esta vía comienza con la desaminación de la fenilalanina y sintetiza 4-cumaroil-CoA por 4CL. Luego, la 4-cumaroil-CoA se convierte en cafeoil-CoA a través de varias reacciones por HCT, que cataliza dos pasos diferentes, seguido de la metilación a través de CCoAOMT y sintetiza feruloil-CoA.1,37. Feruloyl-CoA se convierte en alcohol coniferílico a través de dos reacciones de reducción por CCR y CAD8,38. Dado que el alcohol coniferílico se conoce como un precursor crítico en la biosíntesis de PTOX, los pasos mencionados anteriormente se consideraron aguas arriba.39. En el presente estudio, CAD contiene el mayor número de transcripciones en los pasos anteriores que muestran patrones de expresión espacial y pueden tener diferentes roles funcionales en órganos específicos. Por ejemplo, OsCCR10 implica en respuesta a la sequía en la raíz del arroz40. nocaut de OsCCR10 con el sistema CRISPR/Cas9 reveló que la tolerancia a la sequía redujo el arroz debido a una disminución en el contenido de lignina en la raíz41. Después del alcohol coniferílico, los últimos pasos especificados de PTOX y su biosíntesis derivada se consideraron como un paso posterior. Estos pasos comienzan con el acoplamiento de dos moléculas de alcohol coniferílico para obtener enantiómeros de pinoresinol mediante una oxidasa (LAC11) o una peroxidasa (POD) con la ayuda de proteínas dirigentes, según la especie vegetal.42,43,44. L.usitatissimum genera ambos enantiómeros (-)- y (+)-pinoresinol, seguido de la conversión reductora paso a paso a lariciresinol y luego SECO a través de PLR (1–4)45. (-) SECO, resultante de (+)-pinoresinol, es catalizado por la acción de SDH a matairesinol. Posteriormente, el matairesinol se convierte en desoxipodofilotoxina mediante varias reacciones enzimáticas en P. hexandrum, como PhOMT3, CYP71CU1, PhOMT1 y 2-ODD. Sin embargo, los genes que codifican las enzimas asociadas con los pasos entre matairesinol y desoxipodofilotoxina aún no se han identificado en Linum8,46,47. Este estudio se dio cuenta de algunos de estos genes. El análisis transcriptómico de L.usitatissimum reveló que los niveles más altos de expresión de ALC11, POD, 4CLy Extensión SDH genes estaban bajo estrés por sequía. Las raíces sometidas a estrés abiótico demostraron la regulación a la baja del nivel de expresión de estas enzimas. Por el contrario, la expresión cuantitativa de Extensión SDH usando qRT-PCR mostró una tendencia creciente en L álbum raíces y diferían en sus brotes bajo todos los tratamientos. Además, 2 impares, excluyendo el estrés por sequía, y OMT1 mostró una tendencia creciente en L álbum bajo todo estrés, de acuerdo con estudios previamente informados en P. hexandrum. Sin embargo, esta vía evolucionó de forma independiente en las dos especies.20. Expresión cuantitativa de genes relacionados con la biosíntesis de PTOX, estudiada en diferentes órganos de P. hexandrum, expuso que Extensión SDH, CAD, ccr, y cinamato 4-hidroxilasa los genes aumentaron en los rizomas más que en las raíces37. El aumento de las expresiones génicas relacionadas con el crecimiento y el desarrollo y la biosíntesis de PTOX también se informó a 15 °C en S. hexandrum. Mientras que las expresiones génicas y el contenido de PTOX disminuyeron y los genes que respondían al estrés dominaron a 25 °C en esta planta1. En el estudio realizado por Kumari et al. (2022), genes de fenilalanina amoniaco-liasa (ShPAL), Sh4CL, ShC3H, SHCCoAOMT, ShCOMT, ShCAD, ShDPO, ShPLRy ShSDH regulado al alza, así como un mayor contenido de PTOX en la raíz bajo estrés por sequía. Sin embargo, no hubo evidencia de PTOX en la hoja.35.
El análisis de cromatografía líquida de alto rendimiento (HPLC), consistente con los resultados de qRT-PCR, demostró un aumento del contenido de SECO y la regulación positiva de tres genes seleccionados (Extensión SDH, OMT1y 2 impares) condujo a producir el PTOX más alto en las raíces después de 48 h de tratamiento combinado. Al mismo tiempo, los brotes produjeron PTOX al consumir el precursor de SECO. Bajo estrés de sequía en la raíz a las 12 h, una gran cantidad de SECO y sobreexpresión de 2 impares provocó la conversión de 6-MPTOX y PTOX durante reacciones paso a paso1. Sin embargo, el contenido de PTOX disminuyó debido a la regulación a la baja de 2 impares a las 48 h, según qRT-PCR. Otras enzimas relacionadas con la biosíntesis de lignanos están probablemente involucradas en la generación de 6-MPTOX, por lo que la PTOX disminuyó bajo estrés por sequía. A pesar de la alta cantidad de SECO y la sobreexpresión de 2 impares (en el brote después de 48 h), PTOX disminuyó significativamente debido a la regulación a la baja de Extensión SDH y OMT1. La K+ la deficiencia y el estrés por sequía a menudo tenían patrones de acumulación opuestos de tres lignanos seleccionados. Las diferentes respuestas a las tensiones individuales y combinadas en L álbum se sugiere el complejo mecanismo de regulación de la biosíntesis de lignanos, que requiere mayor investigación. la expresión de PLR in S. hexandrum contradice el contenido de PTOX en los diferentes órganos, proponiendo que el tejido productor de PTOX no necesariamente puede ser su reserva48.
Aplicación de elicitores abióticos y bióticos, incluidos quitosano, jasmonato de metilo, ácido salicílico, extracto de levadura y Ag+, ha establecido principalmente que la expresión de genes asociados con la biosíntesis de lignanos y el contenido de lignanos mejoró en Linum especies in vitro49,50,51,52. Asimismo, un estudio realizado en diferentes accesiones de L álbum bajo estrés por sequía presentó diferentes patrones basados en respuestas fisiológicas y bioquímicas53.
En general, los resultados de los análisis qRT-PCR y HPLC con análisis transcriptómico son consistentes y corroboran estudios previos. Aunque pueden ser diferentes en algunos casos debido a diferencias de especie o genotipo, tipo de cultivo e intensidad del estrés. La K+ la deficiencia y el estrés por sequía a menudo tenían patrones opuestos de regulación ascendente/descendente, y los efectos sinérgicos negativos suponían muchos gastos para la planta. Mientras que el patrón de regulación ascendente/descendente de fusarium era similar al de K+ deficiencia, el estrés por sequía fue similar al tratamiento con ABA.
Por lo tanto, las estrategias basadas en el transcriptoma para especies que probablemente acumulan lignanos ayudarían a identificar características comunes entre especies y señales ambientales para aclarar la ruta de biosíntesis de PTOX.
- Distribución de relaciones públicas y contenido potenciado por SEO. Consiga amplificado hoy.
- PlatoAiStream. Inteligencia de datos Web3. Conocimiento amplificado. Accede Aquí.
- Acuñando el futuro con Adryenn Ashley. Accede Aquí.
- Compra y Vende Acciones en Empresas PRE-IPO con PREIPO®. Accede Aquí.
- Fuente: https://www.nature.com/articles/s41598-023-36102-7

