Expresión de un transportador de ácido dicarboxílico y eliminación de vías de subproductos.
Introducción previa de la vía rTCA en yo orientalis (cepa SA) (Fig. 1B) permitió la producción de SA con un título de 11.6 g/L en fermentaciones en matraz agitado13. Para mejorar aún más el título, intentamos expresar un transportador para SA. MAE1 en Schizosaccharomyces pombe (SpMAE1) resultó ser el transportador de ácido dicarboxílico más eficiente para la exportación de SA18. El codon optimizado SpMAE1 se integró en el genoma de la cepa SA, dando como resultado la cepa SA/MAE1. Se evaluó la producción de SA de las cepas SA y SA/MAE1 mediante fermentaciones en matraz agitado en medios mínimos (SC-URA) con 50 g/L de glucosa en condiciones de oxígeno limitado. La introducción de SpMAE1 mejoró enormemente el título de SA de 6.8 g/l a 24.1 g/l (Fig. 2, Fig. Suplementaria 1Ay figura complementaria. 1B).
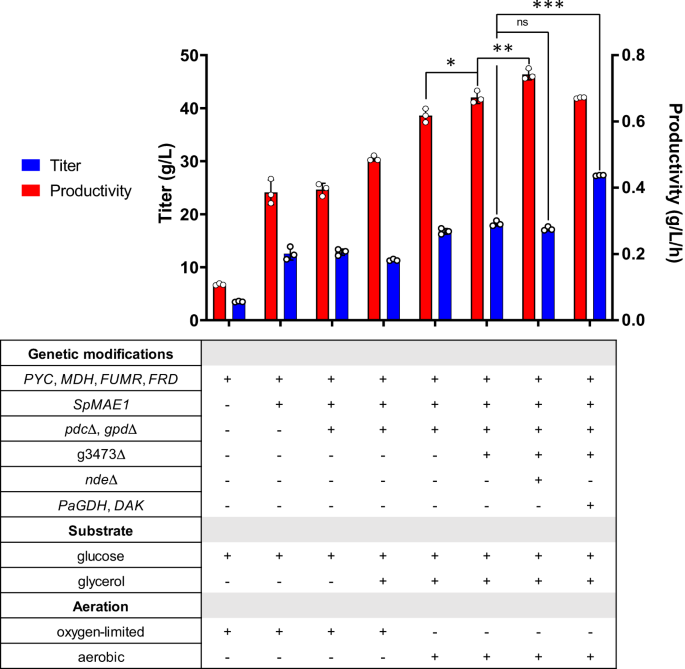
Barras de error, media ± DE (n = 3 muestras biológicamente independientes). *p < 0.05, **p < 0.01, ***p < 0.001, ns no significativo, calculado mediante métodos no apareados de dos colas t-prueba. Los datos de origen se proporcionan como un archivo de datos de origen.
El etanol fue el principal subproducto y se acumuló a 9.5 g/L en la fermentación de la cepa SA/MAE1. El etanol se forma mediante la reacción catalizada por la alcohol deshidrogenasa (ADH), que utiliza NADH para reducir el acetaldehído a etanol. Dado que la producción de SA por la vía rTCA requiere NADH, la eliminación de la vía de formación de etanol podría mejorar la producción de SA. Además, aunque el glicerol se acumuló menos de 1 g/l y no fue el principal subproducto observado en la fermentación de la cepa SA/MAE1, la vía de formación de glicerol catalizada por la glicerol-3-fosfato deshidrogenasa (GPD) puede potencialmente competir con la vía rTCA por carbono y NADH. Así, ambos PDC (piruvato descarboxilasa) y GPD se eliminaron en la cepa SA/MAE1, dando como resultado la cepa SA/MAE1/pdc∆/gpd∆. Mientras que la eliminación de PDC para prevenir la formación de etanol debería teóricamente mejorar el título de SA debido al aumento en la disponibilidad de piruvato y NADH, la fermentación de la cepa SA/MAE1/pdc∆/gpd∆ resultó inesperadamente en un título de SA similar de 24.6 g/L y la acumulación de 19.8 g/L de piruvato en condiciones de oxígeno limitado (Fig. 2 y Fig. Suplementaria. 1C).
Recientemente, se construyó un modelo a escala genómica para yo orientalis, y se predijo que todas las actividades de ADH estarían localizadas en la mitocondria19. Así, la eliminación de la producción de etanol a través de la PDC la eliminación no debería mejorar la disponibilidad de NADH citosólico, lo que no produciría ningún aumento en el título de SA. En cuanto al equilibrio del NADH citosólico, la glucólisis produce 2 moles de piruvato y 2 moles de NADH a partir de 1 mol de glucosa, mientras que la conversión de 1 mol de piruvato en 1 mol de ácido succínico requiere 2 moles de NADH. Por lo tanto, aunque el ciclo reductor de TCA tiene el rendimiento teórico más alto, el rendimiento real de SA en levaduras se limita a sólo 1 mol/mol de glucosa. Realizamos 13C análisis de flujo metabólico (MFA) y verificó que la vía rTCA utilizó eficientemente la mayor parte del NADH citosólico producido por la glucólisis para la producción de SA y la excreción de piruvato representó la mitad del piruvato producido en el último paso de la glucólisis (Figura complementaria. 2). También expresamos una copia adicional de la vía rTCA o gen individual de la vía en la cepa SA/MAE1/pdc∆/gpd∆ para modular aún más los flujos de carbono entre piruvato y SA, pero no hubo cambios significativos en el título de SA o la acumulación de piruvato. (Figura complementaria. 3). Por tanto, la escasez de suministro de NADH en el citosol es el principal cuello de botella para la producción de SA a través de la vía rTCA.
Cofermentación de glucosa y glicerol para la producción de SA.
Dado que la glucosa por sí sola no produce suficiente NADH citosólico para la producción de SA, se pueden considerar otras fuentes de carbono para obtener títulos y rendimientos más altos. El glicerol tiene un mayor grado de reducción y por lo tanto puede producir más equivalentes reductores de NADH que la glucosa.20,21. Dado que la cepa SA/MAE1/pdc∆/gpd∆ presentó defectos de crecimiento en medio SC-URA con glicerol como única fuente de carbono, buscamos realizar la fermentación de esta cepa utilizando medio SC-URA con 50 g/L de glucosa y 20 g /L de glicerol. Anteriormente, se demostró que el uso de glucosa y glicerol como fuentes duales de carbono mejoraba la conversión de oxalacetato en malato mediante el aumento del suministro de NADH a partir de glicerol en un sistema diseñado. M. succinici producens6. Como se muestra en la figura complementaria. 4A, las células podrían consumir ambos sustratos para la producción de SA en condiciones de limitación de oxígeno; sin embargo, el consumo de glicerol fue lento y el título de SA mejoró a sólo 30.5 g/l después de 7 días de fermentación. Luego realizamos la fermentación en condiciones aeróbicas, postulando que el metabolismo del glicerol podría estar limitado en condiciones de oxígeno limitado. En condiciones aeróbicas, tanto la glucosa como el glicerol se consumieron más rápido, lo que permitió la producción de 38.6 g/L de SA (Fig. 2 y Fig. Suplementaria. 4B). También probamos la fermentación de la cepa SA/MAE1/pdc∆/gpd∆ utilizando medio SC-URA con 50 g/L de glucosa en condiciones aeróbicas. Curiosamente, si bien la vía rTCA es una vía fermentativa y una mayor aireación podría canalizar más flujo de carbono hacia el ciclo de TCA para la respiración aeróbica, observamos que las condiciones aeróbicas condujeron a títulos similares en comparación con las condiciones limitadas de oxígeno y las células pudieron consumir glucosa. más rápido, lo que conduce a mayores productividades (Figura complementaria. 4C).
Eliminaciones de un importador de ácido dicarboxílico y NADH deshidrogenasa externa.
Luego se intentaron más eliminaciones de genes para aumentar la producción de SA. Recientemente, los transportadores de carboxilato de la familia JEN PkJEN2-1 y PkJEN2-2 en Pichia kudriavzevii se caracterizaron por estar involucrados en la absorción interna de ácidos dicarboxílicos.22,23. PkJEN2-1 y PkJEN2-2 se anotaron como g3473 y g3068 en yo orientalis, respectivamente. g3473 se eliminó de la cepa SA/MAE1/pdc∆/gpd∆, lo que dio lugar a la cepa g3473∆. La fermentación de esta cepa en medio SC-URA con 50 g/L de glucosa y 20 g/L de glicerol mejoró el título de SA a 42.0 g/L (Fig. 2 y Fig. Suplementaria. 5A), lo que sugiere que impedir que SA vuelva a entrar en las células fue beneficioso. g3068 fue eliminado aún más en la cepa g3473∆; sin embargo, observamos que la interrupción de ambos transportadores JEN2 redujo el título de SA a 34.5 g/l y, por lo tanto, no fue beneficiosa (Figura complementaria. 5B). Este resultado fue inconsistente con el informe anterior de que la eliminación de ambos transportadores JEN en P. kudriavzevii CY902 dio como resultado un título de SA más alto que las eliminaciones de un solo gen, lo que podría atribuirse a diferentes antecedentes genéticos. P. kudriavzevii CY902 fue diseñado para producir SA utilizando la vía oxidativa TCA (oTCA) mediante la eliminación del gen de la subunidad del complejo succinato deshidrogenasa. SDH5, mientras que la producción de SA en nuestro diseño yo orientalis SD108 se logró utilizando la vía rTCA. Además, basándose en MFA, una pequeña cantidad de NADH citosólico fue oxidada por la NADH deshidrogenasa mitocondrial externa (NDE), que transporta electrones desde el NADH citosólico a la cadena de transporte de electrones mitocondrial (Figura complementaria). 2). ECM fue objeto de alteración en la cepa g3473∆, lo que resultó en la cepa g3473∆/nde∆. En comparación con la cepa g3473∆, ECM La eliminación mejoró aún más el título de SA a 46.4 g/l, lo que sugiere la eliminación de ECM aumentó la reserva citosólica de NADH para la producción de SA (Fig. 2 y Fig. Suplementaria. 5C). Sin embargo, la perturbación de ECM redujo la tasa de consumo de glucosa; por lo tanto, a pesar de tener un título más alto, la cepa g3473∆/nde∆ tuvo una productividad similar a la cepa g3473∆ (Fig. 2).
Mejorar el consumo de glicerol
El lento consumo de glicerol indicó que el metabolismo endógeno del glicerol podría no ser muy activo. Anteriormente, la sobreexpresión de glicerol deshidrogenasa (GDH) desde Pichia angusta y dihidroxiacetona quinasa endógena (DAK) estableció una vía de consumo de glicerol productora de NADH en S. cerevisiae24. Por lo tanto, buscamos emplear una estrategia similar para mejorar el consumo de glicerol en yo orientalis. El codón optimizado. PaGDH y endógeno DAK se sobreexpresaron en las cepas g3473∆ y g3473∆/nde∆, lo que resultó en las cepas g3473∆/PaGDH-DAK y g3473∆/nde∆/PaGDH-DAK, respectivamente. Las fermentaciones de estas cepas en medio SC-URA con 50 g/L de glucosa y 20 g/L de glicerol no condujeron a títulos más altos de SA; g3473∆/PaGDH-DAK y g3473∆/nde∆/PaGDH-DAK produjeron SA con títulos de 41.9 g/L y 46.5 g/L, respectivamente, similares a los títulos logrados por las cepas originales que carecían de la sobreexpresión de PaGDH y DAK (Higo. 2, Fig. Suplementaria 6Ay figura complementaria. 6B). Sin embargo, la sobreexpresión de PaGDH y DAK fue beneficioso para las tasas de utilización de glucosa y glicerol. Las productividades aumentaron de 0.29 a 0.44 g/L/h en la cepa g3473∆/PaGDH-DAK y de 0.28 a 0.32 g/L/h en la cepa g3473∆/nde∆/PaGDH-DAK (Fig. 2 y Fig. Suplementaria. 6C).
La cepa g3473∆/PaGDH-DAK podría producir 25.4 g/L de SA en fermentación usando 50 g/L de glucosa, mientras que se podrían obtener 41.9 g/L de SA a partir de 50 g/L de glucosa y 20 g/L de glicerol (Suplementario). Higo. 7A). Dado que el título de SA de 41.9 g/l también podría lograrse simplemente usando más glucosa inicial en la fermentación usando solo glucosa, uno puede cuestionar las ventajas de usar glucosa y glicerol como fuentes duales de carbono. En términos de carbono equivalente, 1 gramo de glucosa equivale a 1 gramo de glicerol. El uso de 50 g/L de glucosa y 20 g/L de glicerol permitió obtener un rendimiento de SA de 0.60 g/g de equivalente de glucosa, que fue mayor que el rendimiento de 0.51 g/g de glucosa de la fermentación utilizando solo 50 g/L de glucosa (Figura complementaria . 7B). Además, a partir de 70 g/L de glucosa, una concentración equivalente a 50 g/L de glucosa y 20 g/L de glicerol, la cepa g3473∆/PaGDH-DAK podría producir SA con un título de sólo 35.6 g/L y un rendimiento de 0.50. g/g de glucosa (Fig. 7). Por lo tanto, utilizar una mezcla de glucosa y glicerol como fuentes de carbono permitió la producción de SA con títulos más altos y rendimientos más altos que usar una cantidad equivalente de glucosa. También comparamos la producción de SA en la cepa g3473∆/PaGDH-DAK usando 50 g/L de glucosa y diferentes concentraciones de glicerol de 10 g/L, 20 g/L y 30 g/L (Figura complementaria. 8). Como se esperaba, el título de SA aumentó a medida que aumentó la cantidad inicial de glicerol; por lo tanto, una comparación entre títulos no fue significativa. Además, el rendimiento fue más bajo cuando se utilizaron 10 g/L de glicerol. No se observaron diferencias significativas en los rendimientos cuando se utilizaron 20 g/L y 30 g/L de glicerol; sin embargo, la productividad fue mayor cuando se utilizaron 20 g/L de glicerol. Por lo tanto, consideramos 20 g/L de glicerol como la concentración óptima para usar con 50 g/L de glucosa para la producción de SA.
También intentamos aliviar la represión catabólica de la glucosa en el consumo de glicerol mediante la eliminación de una hexoquinasa, que demostró reducir la tasa de fosforilación de la glucosa y permitir la coutilización de glucosa y xilosa en S. cerevisiae25. Mediante el análisis BLAST, se determinaron tres genes potenciales de hexoquinasa (g1398, g2945 y g3837), y solo la eliminación de g3837 en la cepa g3473∆/PaGDH-DAK permitió el consumo simultáneo de glucosa y glicerol (Figura complementaria. 9). Si bien se pudieron lograr títulos de SA similares, la eliminación de g3837 redujo las tasas de consumo de glucosa y glicerol, sin producir ningún aumento en la productividad.
Fermentaciones por lotes alimentados y ampliación a escala
Después de las fermentaciones en matraz agitado, realizamos fermentaciones por lotes alimentados para aumentar el título de SA y evaluar el rendimiento de nuestra cepa diseñada en producción a gran escala. Para explotar la tolerancia superior al pH bajo de yo orientalis, elegimos realizar fermentaciones discontinuas a pH 3. A este pH, aproximadamente el 90% de las especies de SA son SA completamente protonadas, mientras que el 10% restante de las especies son succinato de hidrógeno.26. Primero probamos el rendimiento de la cepa g3473∆/PaGDH-DAK, que se eligió sobre g3473∆/nde∆/PaGDH-DAK debido a su mayor productividad, utilizando medio SC-URA con 50 g/L de glucosa y 20 g/L de glicerol en fermentación discontinua en un biorreactor de mesa con un tamaño de 0.3 L y un volumen de trabajo de 0.1 L en condiciones estáticas de agitación y rociado continuo de O2 y compañía2. Observamos que los títulos (27.1 g/L y 30.7 g/L a 0.333 vvm (volumen por volumen de trabajo por min) de CO2 y 0.667 vvm de CO2, respectivamente) fueron mucho más bajos que los títulos obtenidos en la fermentación en matraz agitado (42.1 g/L) (Fig. 10A,B). En particular, si bien se pudieron producir títulos similares de SA a partir de glucosa tanto en el reactor como en el matraz de agitación, los títulos de SA producidos durante la fase de utilización de glicerol fueron mucho más bajos en el biorreactor. También realizamos una fermentación discontinua en biorreactor utilizando la cepa g3473∆/PaGDH-DAK/g3837∆ y observamos que esta cepa podía producir más SA durante la fase de consumo de glicerol y se podía obtener un título de 38.8 g/L de SA a 0.167 vvm de O.2 y 0.667 vvm de CO2 (Fig. complementaria. 10C,D). Postulamos que mientras se utilizaba glicerol en el ambiente del biorreactor con mayor aireación que el ambiente del matraz agitado, se podría canalizar más flujo de carbono al ciclo de TCA y conducir a un título de SA más bajo; por otro lado, la eliminación de g3837 podría reprimir la actividad de los genes del ciclo TCA y mejorar la producción de SA. Se empleó un análisis de PCR en tiempo real para comparar los niveles transcripcionales de genes en la vía rTCA y algunos genes seleccionados en el ciclo TCA (citrato sintasa, CIT; aconitasa, ACO; e isocitrato deshidrogenasa, HDI) en cepas g3473∆/PaGDH-DAK con o sin deleción g3837 cultivadas en medio YP con glicerol. Observamos que la eliminación de g3837 mantuvo expresiones similares de genes en la vía rTCA pero redujo los niveles de expresión de CIT, un homólogo de ACOy HDIs (Figura complementaria. 11). Por lo tanto, las menores actividades de los genes en el ciclo de TCA podrían conducir a un mayor título de SA obtenido por la cepa g3473∆/PaGDH-DAK/g3837∆ en el biorreactor. La fermentación discontinua alimentada de la cepa g3473∆/PaGDH-DAK/g3837∆ en medio SC-URA con alimentación de glucosa y glicerol produjo 109.5 g/L de SA con un rendimiento de 0.65 g/g de equivalente de glucosa y una productividad de 0.54 g/ L/h (Fig. 3A). Al final de la fermentación observamos la formación de cristales, probablemente de SA (Fig. 12). Mientras que otros ácidos orgánicos, como los ácidos láctico y acético, son completamente miscibles en caldo acuoso a un pH de 1 a 14, la solubilidad del SA disminuye a medida que el pH se vuelve más ácido.27.
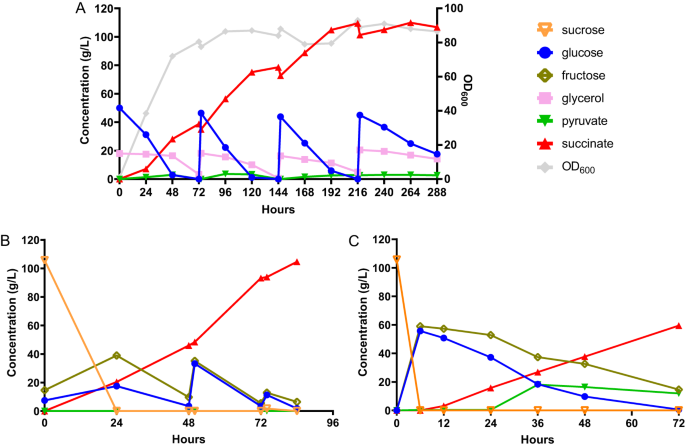
A Fermentación por lotes alimentados de la cepa g3473∆/PaGDH-DAK/g3873∆ en medio mínimo con glucosa y glicerol. B Fermentación por lotes alimentados de la cepa g3473∆/PaGDH-DAK/ScSUC2 en medio de jugo de caña de azúcar. C Fermentación discontinua de la cepa g3473∆/PaGDH-DAK/ScSUC2 en medio de jugo de caña de azúcar en un reactor a escala piloto. Los datos de origen se proporcionan como un archivo de datos de origen.
Tras el alto rendimiento fermentativo de nuestro recombinante yo orientalis cepa utilizando el medio mínimo comúnmente utilizado en el laboratorio, luego probamos la producción de SA utilizando un sustrato industrial real, el jugo de caña de azúcar. La caña de azúcar es la planta perenne C4 con mayor eficiencia energética y tiene un mayor rendimiento de biomasa en comparación con otros cultivos como el pasto varilla y el miscanto.28. Además, el jugo de caña de azúcar, como materia prima a base de sacarosa, es más barato que los sustratos a base de glucosa y almidón, como el maíz y la yuca.29. Desde yo orientalis no puede utilizar sacarosa, la invertasa SUC2 en S. cerevisiae se expresó en g3473∆/PaGDH-DAK. La fermentación discontinua se produjo en las primeras 48 h y se pudo producir SA con un título de 46.0 g/l, un rendimiento de 0.40 g/g de equivalente de glucosa y una productividad de 0.96 g/l/h. Con la alimentación posterior con jugo de caña de azúcar concentrado, nuestra cepa diseñada podría producir SA a un título de 104.6 g/L, un rendimiento de 0.63 g/g de equivalente de glucosa y una productividad de 1.25 g/L/h a escala de laboratorio (Fig. 3B).
Además, ampliamos nuestro proceso de fermentación SA utilizando jugo de caña de azúcar desde la escala de banco a una escala piloto. Aquí, determinamos los parámetros del proceso para mantener una entrada de energía similar por unidad de volumen y el mismo número de Reynolds entre los biorreactores a escala de banco y a escala piloto, y la fermentación por lotes se realizó en un biorreactor a escala piloto con un tamaño de 75 L y una capacidad de trabajo. volumen de 30 L o un factor de ampliación de 300 × en comparación con el biorreactor de mesa. Nuestra cepa podría producir SA con un título de 63.1 g/L, un rendimiento de 0.50 g/g de equivalente de glucosa y una productividad de 0.66 g/L/h a pH 3 (Fig. 3C). Debido al requisito de volumen, no intentamos una fermentación discontinua a escala piloto; sin embargo, nuestro título y rendimiento para la fermentación discontinua en el biorreactor a escala piloto fueron comparables a los de los biorreactores a escala de laboratorio. Por lo tanto, anticipamos un desempeño similar de la cepa en la fermentación por lotes alimentados a escala piloto.
Además, completamos todo el proceso de producción de SA mediante el diseño de un DSP para recuperar SA del caldo de fermentación del jugo de caña de azúcar mediante destilación al vacío y cristalización en dos etapas. Sin acidificación adicional del caldo de fermentación que contenía 63.1 g/L de SA obtenido de la fermentación a escala piloto, el rendimiento máximo fue del 31.0 % durante la primera etapa. Luego se concentró el filtrado de la etapa 1 hasta el 50% de su volumen mediante destilación al vacío y se sometió a la segunda etapa de cristalización. El rendimiento de SA de la etapa 2 fue del 47.7% y se obtuvieron cantidades similares de cristales de SA para ambas etapas (1.98 g para la etapa 1 y 2.10 g para la etapa 2). Así, mediante destilación al vacío y cristalización en dos etapas, se obtuvo un rendimiento global de recuperación de SA del 64.0 % a partir del caldo de fermentación de bajo pH. Además, se estimó que las purezas de los cristales de SA recuperados en las etapas 1 y 2 eran 88.9% y 86.23%, respectivamente. El resultado obtenido estuvo en línea con los hallazgos de un estudio anterior.30. A partir de los resultados, fue evidente que la cristalización de SA con alta pureza (>85%) a partir del caldo de fermentación sin tratar fue exitosa. Sin embargo, se necesita más investigación para eliminar las impurezas colorantes para recuperar el cristal SA de calidad comercial.
Análisis tecnoeconómico y evaluación del ciclo de vida.
Diseñamos y simulamos biorrefinerías de extremo a extremo capaces de aceptar caña de azúcar como materia prima, sacarificarla hasta obtener jugo de caña de azúcar (sacarosa, glucosa y fructosa), fermentar los azúcares hasta obtener SA utilizando yo orientalis, y separar el caldo de fermentación para recuperar los cristales de SA secos (Fig. 13) con una capacidad de producción anual de 26,800 toneladas métricas de SA (la demanda global de SA en 2013 fue de aproximadamente 76,000 toneladas métricas31). Las biorrefinerías se simularon en escenarios de fermentación alternativos con supuestos de rendimiento, título y productividad correspondientes al rendimiento de fermentación logrado en experimentos en modo discontinuo a escala de laboratorio (escenario discontinuo de laboratorio) y en modo discontinuo alimentado (escenario discontinuo de laboratorio), así como la configuración del modo por lotes a escala piloto (escenario por lotes piloto). Para caracterizar la viabilidad financiera y los beneficios ambientales de las rutas de SA desarrolladas, realizamos TEA y LCA para cada escenario bajo supuestos de referencia así como bajo incertidumbre (simulaciones de Monte Carlo de 2000 para cada escenario con muestreo de hipercubo latino; la distribución de resultados en números alternativos de las simulaciones de Monte Carlo se informa en la tabla complementaria 1 y los valores de referencia supuestos y las distribuciones de todos los parámetros inciertos para cada escenario se informan en Datos complementarios. 1). Utilizamos el precio mínimo de venta del producto (MPSP, en 2016$ con una tasa interna de retorno del 10%), potencial de calentamiento global a 100 años (GWP100; cradle-to-grave) y el consumo de energía fósil (FEC; cradle-to-gate) como métricas para representar los resultados de TEA y LCA. También realizamos análisis de sensibilidad utilizando los coeficientes de correlación de orden de rango de Spearman (ρ de Spearman) para identificar los factores clave de los costos de producción y los impactos ambientales. Finalmente, para establecer y priorizar objetivos para mejorar aún más la viabilidad financiera y la sostenibilidad ambiental, diseñamos y simulamos biorrefinerías en todo el panorama potencial de rendimiento de fermentación (es decir, 2500 combinaciones de rendimiento-título cada una en una gama de productividades para fermentación neutra y de bajo pH). ).
Con base en el desempeño experimental en el escenario del lote de laboratorio, la biorrefinería podría producir SA a un MPSP estimado de $1.70/kg (línea de base; Fig. 4A) con un rango de 1.51 a 1.92 dólares/kg (percentiles 5 a 95; en adelante entre paréntesis). El PCA de la biorrefinería100 y el FEC en este escenario se estimó en 1.95 kg de CO2-eq./kg (1.37–2.65 kg CO2-eq./kg) y -3.74 MJ/kg (−12.9–5.39 MJ/kg), respectivamente (Fig. 4B y Fig. Suplementaria. 14A). En el escenario de laboratorio alimentado por lotes (con títulos de SA de fermentación, rendimiento y productividad mejorados respecto a los del escenario de laboratorio por lotes), el MPSP de la biorrefinería fue de $1.06/kg ($0.96–1.22/kg), GWP100 fue de 0.93 kg de CO2-eq./kg (0.71–1.32 kg CO2-eq./kg), y la FEC fue −5.36 MJ/kg (−8.97–0.213 MJ/kg). En el escenario del lote piloto (con rendimiento y título de SA de fermentación mejorados en relación con el escenario del lote de laboratorio pero menores que los del escenario de lote alimentado de laboratorio, y menor productividad que ambos escenarios de laboratorio), la biorrefinería tenía un MPSP estimado de $1.37/kg. ($1.23–1.54/kg), PCA100 de 1.67 kg CO2-eq./kg (1.22–2.17 kg CO2-eq./kg) y FEC de −0.21 MJ/kg (−7.08 a 6.47 MJ/kg). En la figura complementaria también se muestra un diagrama de Sankey que representa el flujo de carbono a través de la biorrefinería para este escenario. 15.
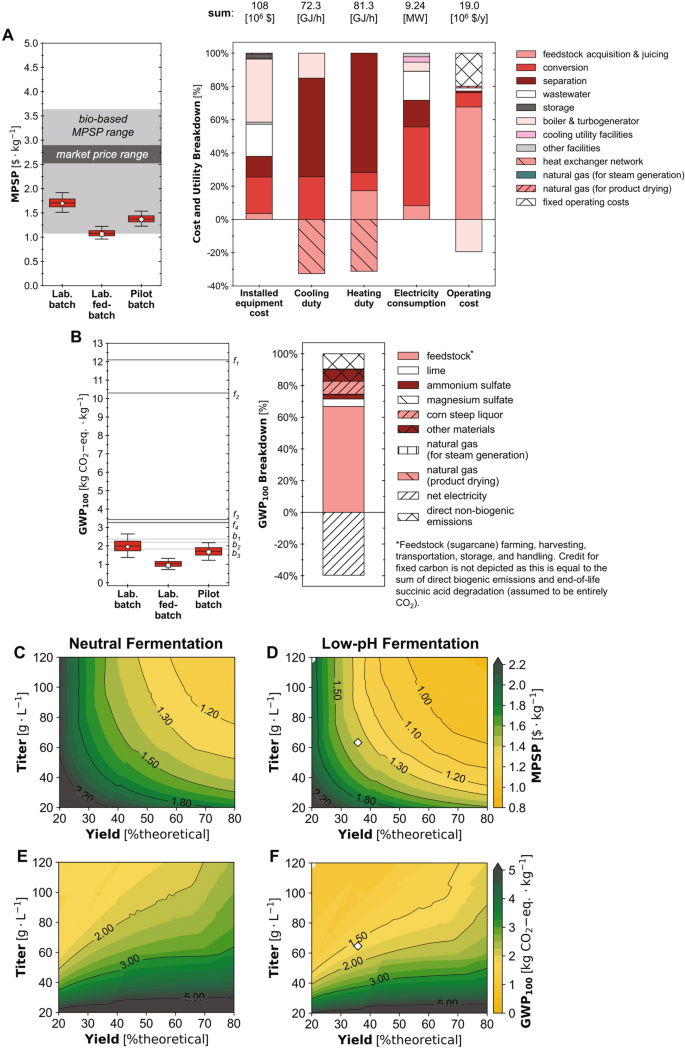
Incertidumbres (gráficos de cajas y bigotes) y desgloses (gráficos de barras apiladas) para (A) precio mínimo de venta del producto (MPSP) y (B) potencial de calentamiento global (GWP) desde la cuna hasta la tumba en 100 años100). Los bigotes, las cajas y la línea media representan los percentiles 5/95, 25/75 y 50 de simulaciones de Monte Carlo de 2000 (n = 2000 simulaciones) para cada escenario. Los marcadores de pentágono, cuadrado y diamante representan resultados de referencia para los escenarios de lote de laboratorio (Lab. lote), lote alimentado de laboratorio (Lab. lote alimentado) y lote piloto (lote piloto), respectivamente. Los gráficos de barras apiladas informan los resultados de referencia para el escenario del lote piloto; Los resultados para otros escenarios se incluyen en el SI. El consumo de electricidad incluye únicamente el consumo del sistema; la producción se excluyó en el desglose representado para mayor claridad de la figura. Datos desglosados tabulados para costos de materiales y equipos instalados, tareas de calefacción y refrigeración, uso de electricidad, GWP100y FEC están disponibles en línea62. Las líneas marcadas en gris oscuro indican los impactos reportados para las vías de producción basadas en fósiles (f152; f2-f451). Las líneas marcadas en gris claro denotan los impactos reportados para vías alternativas de producción de base biológica (b151; b253; b352). Donde GWP100 se informó como de la cuna a la puerta, se agregaron 1.49 kg CO2-eq./kg como impactos al final de la vida para mantener la coherencia con este estudio y Dunn et al. 2015. Los valores de todos los MPSP informados y los impactos antes y después del ajuste se enumeran en la Tabla complementaria 3 y 4 (C, D) MPSP y (E, F) PCA100 en 2500 combinaciones de rendimiento de fermentación y título en la productividad inicial del escenario del lote piloto (0.66 g/L/h) para neutral (panel izquierdo; C, E) y pH bajo (panel derecho; D, F) fermentación. El rendimiento se muestra como % del máximo teórico (%teórico) escalado al rendimiento máximo teórico de 1.31 g/g de equivalente de glucosa (basado en el equilibrio de carbono). Para un punto dado de la figura, la xEl valor del eje -representa el rendimiento, el valor del eje y representa el título y el color y las líneas de contorno representan el valor de MPSP, GWP100. Los marcadores de diamantes muestran los resultados de referencia para el escenario del lote piloto. Los datos de origen se proporcionan como un archivo de datos de origen.
Entre los 28 parámetros a los que se atribuyó la incertidumbre para el escenario del lote piloto, encontramos que el MPSP era más sensible al rendimiento de SA de fermentación (ρ de Spearman de −0.60; todas las distribuciones de incertidumbre se enumeran en Datos complementarios 1 y los valores ρ de Spearman para todos los parámetros se informan en la tabla complementaria 2). MPSP también fue significativamente sensible al precio unitario de la caña de azúcar forrajera (ρ de Spearman de 0.39), el tiempo de actividad de la planta (-0.38), la capacidad de la planta para alimentar la caña de azúcar (-0.31) y el título de SA de fermentación (-0.30). PCA100 fue más sensible a la eficiencia de la caldera (ρ de Spearman de −0.63). PCA100 también fue significativamente sensible al título y al rendimiento de SA de fermentación, con valores de ρ de Spearman de −0.62 y 0.32, respectivamente. La FEC fue más sensible al rendimiento de SA de fermentación (ρ de Spearman de 0.63), y también sensible a la eficiencia de la caldera (-0.56) y al título de SA de fermentación (-0.49). Para caracterizar mejor las implicaciones del rendimiento de la fermentación, realizamos TEA y ACV en todo el panorama del rendimiento de la fermentación (Fig. 4C – F y Fig. Suplementaria. 14B, C), simulando 2500 combinaciones de rendimiento-título en una gama de productividades para cada uno de dos regímenes alternativos: fermentación con pH bajo (es decir, con fermentación a un pH de 3 controlado mediante la adición de base durante la fermentación y sin acidulación requerida después de la fermentación) y fermentación neutra. (es decir, con neutralización completa de SA mediante adición de base durante la fermentación y reacidulación completa después de la fermentación). Los resultados para un escenario alternativo de pH bajo con reacidulación requerida después de la fermentación se muestran en la figura complementaria. 16.
- Distribución de relaciones públicas y contenido potenciado por SEO. Consiga amplificado hoy.
- PlatoData.Network Vertical Generativo Ai. Empodérate. Accede Aquí.
- PlatoAiStream. Inteligencia Web3. Conocimiento amplificado. Accede Aquí.
- PlatoESG. Carbón, tecnología limpia, Energía, Ambiente, Solar, Gestión de residuos. Accede Aquí.
- PlatoSalud. Inteligencia en Biotecnología y Ensayos Clínicos. Accede Aquí.
- Fuente: https://www.nature.com/articles/s41467-023-41616-9

